 ?萍加邢薰? />
?萍加邢薰? />
湖北省益境環(huán)?div id="jpandex" class="focus-wrap mb20 cf">?萍加邢薰?!-- 益境環(huán)?div id="jpandex" class="focus-wrap mb20 cf">?萍?font style="font-size:22px;">-專業(yè)從事各種廢水處理及回用工程 -->
專業(yè)從事煙氣除塵
全國服務(wù)熱線
湖北省益境環(huán)?div id="jfovm50" class="index-wrap">?萍加邢薰?/strong>
地址:湖北省大冶市羅橋工業(yè)園大冶大道284號
電話:0714-8758589
13797788774
13117030666
傳真:0714-8758579
郵編:435000
網(wǎng)址:http://m.tywork.cc
詳細(xì)內(nèi)容
酸性礦山廢水氨氧化菌群落研究
大規(guī)模金屬礦藏的開采
低pH和高金屬含量不僅影響周圍地區(qū)的生態(tài)系統(tǒng)
本研究取樣點位于安徽省某酸性礦山廢水區(qū)域,采集有短小植被覆蓋區(qū)域的根際土壤
1 材料與方法
1.1 樣品采集及分析
樣品于2012年8月采集于安徽某鐵礦酸性礦山廢水庫邊有短小植被覆蓋區(qū)域. 采集根際土壤樣品后,裝于無菌封口袋中
1.2 土壤基因組DNA的提取
取0.5 g土壤樣品
1.3 細(xì)菌16S rRNA基因的擴增
利用細(xì)菌通用引物27F(5′-AGA GTT TGA TCM TGG CTC AG-3′)和1492R(5′-TAC GGY TAC CTT GTT ACG ACT T-3′)[17]
1.4 古菌16S rRNA基因的擴增
以土壤基因組DNA為模板
1.5 氨單加氧酶基因(amoA)的擴增
amoA是氨氧化功能酶——氨單加氧酶(ammonia monooxygenase,AMO)α亞基的編碼基因. 以土壤基因組DNA模板,分別利用氨氧化細(xì)菌amoA引物amoA-1F(GGG GHT TYT ACT GGT GGT)和amoA-2R(CCC CTC KGS AAA GCC TTC TTC)[19]
、 amoA-3F(GGT GAG TGG GYT AAC MG)和amoA-4R(GCT AGC CAC TTT CTG G)[20],氨氧化古菌amoA引物AF(STA ATG GTC TGG CTT AGA)和AR(GCG GCC ATC CAT CTG TAT GT)[21]擴增樣品中氨氧化細(xì)菌和氨氧化古菌的氨單加氧酶基因片段. 氨氧化細(xì)菌PCR反應(yīng)條件設(shè)置為95℃ 5 min,變性94℃ 30 s,退火48℃ 45 s,延伸72℃ 2 min,35個循環(huán)后,72℃延伸10 min. 氨氧化古菌PCR反應(yīng)條件設(shè)置為95℃ 5 min,變性94℃ 30 s,退火48℃ 60 s,延伸72℃ 2 min,35個循環(huán)后,72℃延伸10 min.1.6 克隆文庫的構(gòu)建
分別構(gòu)建土壤樣品中細(xì)菌和古細(xì)菌的16S rDNA克隆文庫以及氨氧化古菌的amoA克隆文庫. 氨氧化細(xì)菌amoA基因經(jīng)反復(fù)擴增
,均未得到PCR產(chǎn)物,故未構(gòu)建該文庫. 克隆文庫構(gòu)建方法為:將PCR產(chǎn)物連接到Promega公司的pGEM-T克隆載體上1.7 序列測定以及系統(tǒng)發(fā)育分析
利用Bellerophon程序檢測并去除嵌合體[22]. 然后用Dotur軟件對所得序列進行分類
1.8 序列登錄號
將本研究中所得序列提交至GenBank中
,細(xì)菌16S rDNA的序列登錄號為KF863913-KF863952,KF924200-KF924205; 古菌16S rDNA的序列登錄號為KF918278-KF918306; 氨氧化古菌amoA的序列登錄號為KF916687-KF916696.2 結(jié)果與分析
2.1 AODC結(jié)果與理化分析
所采集土壤樣品的部分物理化學(xué)參數(shù)見表 1. 樣品酸性很強
,pHKCl僅為2.83. 總有機碳含量為0.675%,低于農(nóng)田和森林土壤TOC平均含量[23,24]; 含水率為9.5%; 土壤微生物豐度為3.1×108 個 ·g-1; NH+4-N和NO-3-N含量分別為7.6 mg ·kg-1、 3.1 mg ·kg-1. 經(jīng)測得樣品中有效金屬含量很高,其中Al含量最高達1 380.3 mg ·kg-1,F(xiàn)e含量高達57.0 mg ·kg-1. SO2-4含量高達2 168.0 mg ·kg-1.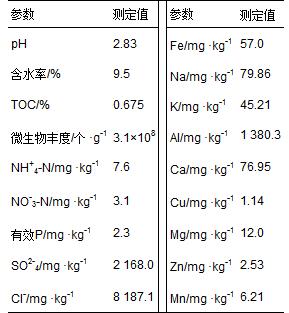
表 1 土壤樣品的部分化學(xué)參數(shù)
2.2 飽和度及多樣性指數(shù)分析
經(jīng)測序細(xì)菌文庫共獲得103條序列
,去除載體和嵌合體后有效序列為95條; 古細(xì)菌與氨氧化古菌文庫分別得到28條和39條有效序列. 3個文庫的飽和曲線如圖 1所示,3個文庫飽和曲線均趨于平緩,可以比較完整地反映樣品中細(xì)菌、 古菌和氨氧化古菌群落結(jié)構(gòu). 從香農(nóng)指數(shù)(Shannon)和辛普森指數(shù)(Simpson)可以看出該樣品中細(xì)菌的多樣性高于古菌(表 2).
圖1 細(xì)菌
、 古菌和氨氧化古菌飽和度曲線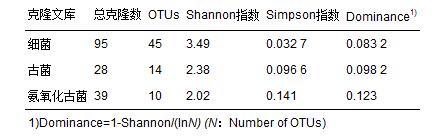
表 2 細(xì)菌
、 古菌以及氨氧化古菌克隆文庫多樣性指數(shù)2.3 土壤中細(xì)菌、 古菌以及氨氧化古菌多樣性以及系統(tǒng)發(fā)育學(xué)分析 2.3.1 細(xì)菌多樣性以及系統(tǒng)發(fā)育分析
將每種基因型的序列輸入RDP網(wǎng)站
,得到該基因型所屬的系統(tǒng)發(fā)育類群. 結(jié)果顯示,細(xì)菌文庫中包含了11個類群(如圖 2),其中所占比例最大的3個類群分別為Acidobacteria(47.4%)、 Verrucomicrobia(18.9%)、 Chloroflexi(10.5%).
圖 2 細(xì)菌文庫中各類群所占比例
經(jīng)NCBI數(shù)據(jù)庫中Blast程序比對分析后
,選取主要克隆進行比對,發(fā)現(xiàn)與該礦區(qū)土壤樣品中細(xì)菌親緣關(guān)系較近的克隆均來自于一些酸性環(huán)境中,如某些礦區(qū)酸性土壤、 酸性濕地底泥、 酸性紅壤和酸性礦山廢水等,或者植被根際等.本研究中酸桿菌門是所占比例最高的門類
,在文庫中豐度達47.4%. 酸桿菌門細(xì)菌在環(huán)境中分布廣泛
圖 3 基于16S rDNA序列的Acidobacteria細(xì)菌系統(tǒng)發(fā)育樹
克隆Bac-XC-105(2.0%)
、 Bac-XC-109(5.1%)、 Bac-XC-19(1.0%)和Bac-XC-25(1.0%)均與Candidatus Koribacter versatilis 的親緣關(guān)系較近. Ward等[31]比較了包括Candidatus Koribacter versatilis在內(nèi)的酸桿菌門中3個種的全基因組,發(fā)現(xiàn)Candidatus Koribacter versatilis具備硝酸鹽和亞硝酸鹽還原能力,且適宜在pH 4.0~6.5的環(huán)境中生存.克隆Bac-XC-27(4.0%)
、 Bac-XC-86(1.0%)和Bac-XC-99(1.0%)與菌株Acidobacteria sp. isolate N3B的16S rDNA序列相似性均為97%,該菌株分離自法國加爾省一流經(jīng)古代礦區(qū)的酸性溪流中[32]. Acidobacteria屬菌株多為分離自酸性環(huán)境的化能異養(yǎng)菌,革蘭氏染色呈陰性,好氧; 可在pH 3.0~6.0、 溫度為20~37℃的環(huán)境中生存[33].克隆Bac-XC-14和Bac-XC-4(豐度分別為4.0%和3.0%)與Holophaga sp.的16S rDNA序列同源性較高. Holophaga屬細(xì)菌在一些酸性礦坑水中曾檢測到
,如江西德興市銀山鉛鋅礦的露天礦坑儲水池和地下300多m隧道中自然氧化形成的酸性礦坑水[34]. Holophaga屬細(xì)菌多為革蘭氏陰性嚴(yán)格厭氧菌,可在溫度為10~35℃環(huán)境中生存,最適溫度為28~32℃,最適pH為6.8~7.5[35].克隆Bac-XC-41(5.1%)和Bac-XC-80(2.0%)在系統(tǒng)發(fā)育樹上與Acidopila rosea聚為一類(圖 3). Okamura等[36]分別在日本的一個AMD處理廠(水樣pH 2.7)和一個茶葉種植園土壤(pH 4.5)中分離到兩株Acidipila rosea. Acidipila rosea為革蘭氏陰性好氧菌
,細(xì)胞呈球形或球桿形,營養(yǎng)類型為化能有機營養(yǎng)型; 可以在pH 3.0~6.0范圍內(nèi)生存,最適pH為4.5; pH 7.0時不能生存.如圖 4所示
,放線菌門包括3種基因型(占文庫的4.0%)都與Aciditerrimonas ferrireducens在系統(tǒng)發(fā)育樹上聚為一簇. Aciditerrimonas ferrireducens是一種中度嗜熱的嗜酸菌,具有還原鐵的能力; 革蘭氏染色呈陽性,可以在pH 2.0~4.5,溫度35~58℃的環(huán)境中生存,最適pH為3.0,最適生長溫度為50℃. 有氧條件下營養(yǎng)類型為化能異養(yǎng),無氧環(huán)境中為化能自養(yǎng),利用三價鐵氧化氫獲得能量[37].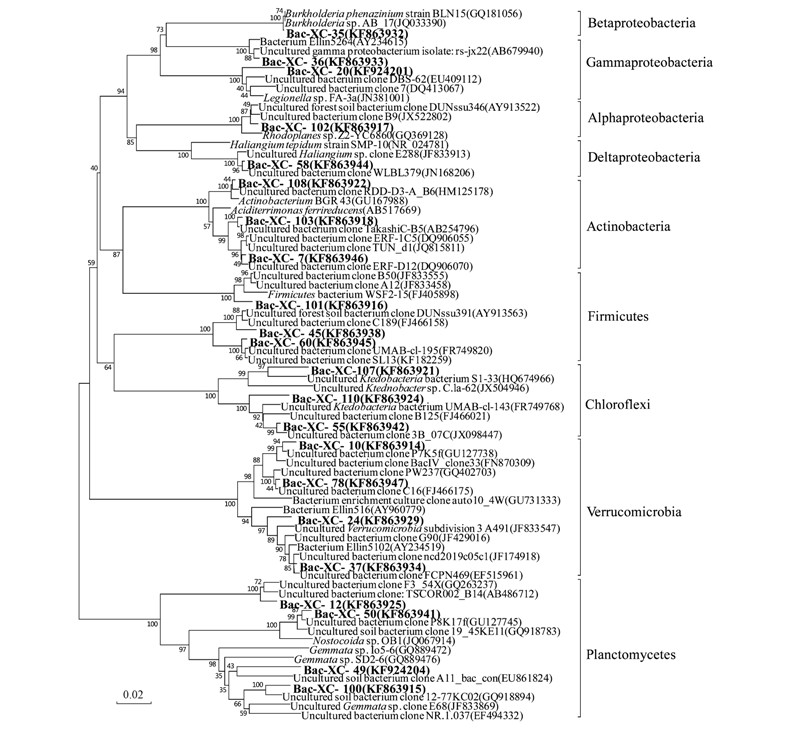
圖 4 基于16S rDNA序列的其它門細(xì)菌系統(tǒng)發(fā)育樹
α-變形菌門在本文庫中所占比例僅為6.0%. 其中克隆Bac-XC-102(3.0%)與紅游動菌屬(Rhodoplanes sp.)Z2-YC6860在系統(tǒng)發(fā)育樹上聚為一簇. 紅游動菌屬細(xì)菌為一類紫色非硫細(xì)菌; 細(xì)胞革蘭氏染色呈陰性
2.3.2 古菌多樣性以及系統(tǒng)發(fā)育分析
經(jīng)過RDP網(wǎng)站的比對分析
奇古菌門是2008年從中溫環(huán)境來源的古菌(溫泉古菌)劃分出來的一個新類群[39]. 古菌文庫中歸類于奇古菌門的古細(xì)菌均與氨氧化作用相關(guān). 奇古菌門的氨氧化古菌分布廣泛
奇古菌門中比例最高也是文庫中豐度最大的基因型為Arch-XC-25,所占比例為27.6%. 在GenBank數(shù)據(jù)庫的比對結(jié)果中
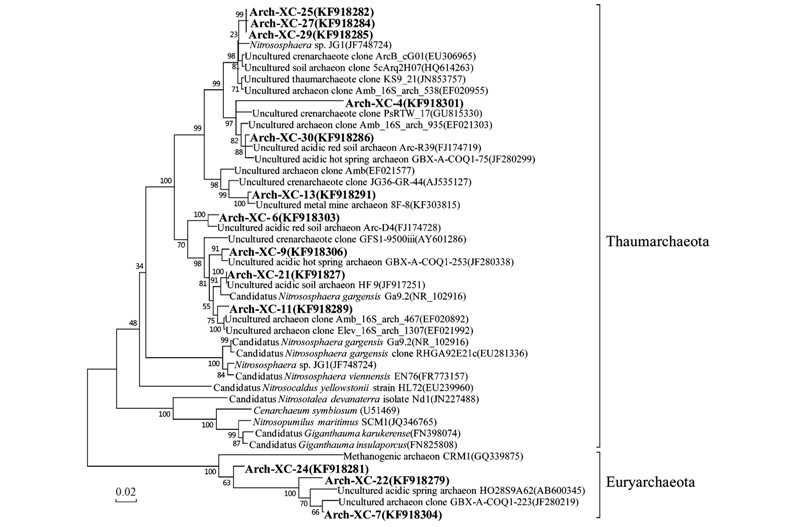
圖 5 基于16S rDNA序列的古菌系統(tǒng)發(fā)育樹
本研究的文庫中廣古菌門包括3種基因型
2.3.3 氨氧化古菌多樣性以及系統(tǒng)發(fā)育分析
該土壤樣品未擴增到氨氧化細(xì)菌的amoA基因片段
由古菌文庫組成可知本樣品中大多數(shù)古菌與氨氧化功能相關(guān),故利用古菌特異性氨單加氧酶引物對AF和AR擴增氨單加氧酶基因序列
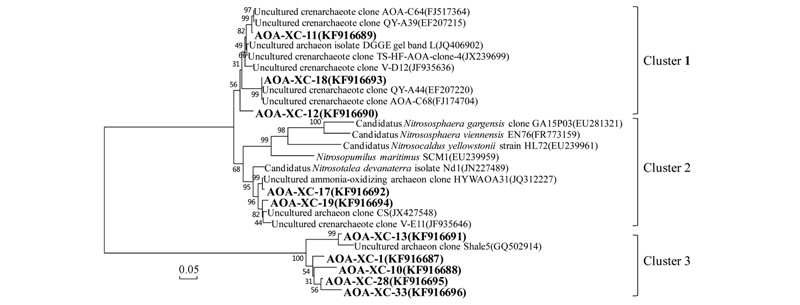
圖 6 基于amoA序列的氨氧化古菌(AOA)系統(tǒng)發(fā)育樹
3 討論
在前期研究中
4 結(jié)論
(1) 取樣區(qū)域土壤受到酸性礦山廢水的嚴(yán)重污染,pH僅為2.83
(2) 細(xì)菌文庫覆蓋11個類群
(3) 細(xì)菌文庫中未發(fā)現(xiàn)與氨氧化作用相關(guān)的類群
資訊
- + 多項目同時開工,益境環(huán)保賣力趕工!
- + 拓展市場積極參與投標(biāo)
- + 純度高達99.99%以上的石英砂成套設(shè)備(可配備污水處理設(shè)備)
- + 南京高淳啟動新一輪雨污分流工程 預(yù)計明年實現(xiàn)雨污分流全覆蓋
- + 聚合氯化鋁如何去除污泥水中的磷
- + 酸性礦山廢水氨氧化菌群落研究
- + FePMo12催化電如何降解染料廢水
- + 樹脂化工集中區(qū)廢水廠尾水的解決方法
- + 玉溪市污水處理廠污泥處理處置工程選址進入初步論證階段
- + 莆田市擬投資1400萬元加快秀嶼區(qū)鎮(zhèn)鎮(zhèn)通污水主管網(wǎng)工程建設(shè)
- + 胞外聚合物對活性污泥吸附生活污水碳源的影響
- + 如何去除制革廢水
- + 連云區(qū)宿城街道污水處理廠建成并投用 規(guī)模高達1000噸/日
- + 陜西已建成120座污水處理廠 規(guī)模高達419.28萬m3/d
地址:湖北省大冶市羅橋工業(yè)園大冶大道284號
電話:0714-8758589 13797788774 13117030666
傳真:0714-8758579 郵編:435000
郵箱:sales@cnyjhb.com 網(wǎng)址:m.tywork.cc
重點關(guān)鍵詞:一體化污水處理設(shè)備
湖北省益境環(huán)保科技有限公司 版權(quán)所有(C)2016 網(wǎng)絡(luò)支持 生意寶 著作權(quán)聲明
鄂ICP備14011879號
 鄂公網(wǎng)安備 42028102000207號
鄂公網(wǎng)安備 42028102000207號

手機版

小程序
| |||||||||